根瘤是一个复杂的共生固氮工厂,其中的细胞是高度异质性的。然而,根瘤细胞的分化和相互联系在很大程度上仍然是未知的。
2022年10月28日,国际权威学术期刊Molecular Plant发表了中国农业大学王涛/董江丽团队的最新相关研究成果,揭示了揭示共生固氮根瘤的单细胞转录组。
近日,法国图卢兹大学Delphine Capela团队在国际著名期刊Nature Plants发表了题为“Comparative phylotranscriptomics reveals ancestral and derived root nodule symbiosis programmes”的文章。
该研究利用比较和系统转录组学来阐明RNS的起源和进化,比较了9种寄主植物的共生转录组反应,重建了由已知的共生基因和数百个新候选基因组成的原始RNS转录组,认为RNS的起源很可能是在9000万年前NFN进化支受到辐射之前演化出来的,并重建了RNS植物共生程序的进化过程。
首先,该研究使用9个物种组成简化系统发育,在树的顶端绘制了每个包含DEG的正构群(以下简称DEOG,为差异表达正构群)的存在/缺失情况,确定哪个进化节点的基因已被RNS吸收,利用固定速率、连续时间及马尔可夫模型来重建离散特征的祖先状态。利用这种方法,确定了在共生过程中基因在哪个系统发育节点上发生了差异调控。其中三个基因(LjNFR5/MtNFP、NIN和RPG)的系统发育分布被认为与NFN进化支中形成RNS的能力有关。事实上,这三个基因已经在多个谱系中独立丢失,不再能够形成RNS。RNS仅在两个谱系中进化稳定,蝶目亚科和蝶目含虫亚科。

NFN物种间RNS转录组反应的保存
通过对模型物种RNS的遗传解剖表明,RNS的进化依赖于调节更古老的AMS和侧根发育基因的共择。为了确定这两个程序对原始RNS转录组反应的贡献,研究交叉引用了模型豆科植物截尾苜蓿(Medicago truncatula)侧根发育和AMS差异调控基因(Lotus japonicus和M. truncatula)与推断的祖先转录组的RNA-seq数据。结果表明RNS的进化主要是通过将较老的侧根和AMS的共同选择结合而进行的。
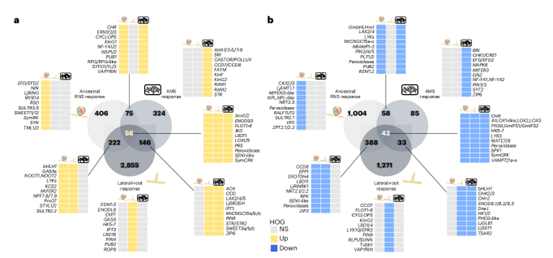
祖先RNS、AMS和侧根转录组中共有的和特异的差异表达
为了定义与每个过程相关的转录模块及其进化起源,该研究利用了一组细菌突变体诱导了完整的共生程序。结果表明hrcV(编码三型分泌系统结构蛋白的基因)中停止突变赋予了细菌结瘤的能力,但结瘤仅被细胞外侵入(图3a)。相比之下,hrpG(一种编码包括三型分泌系统基因在内的数百个基因的整体调控因子)的停止突变使细菌能够形成结节,并通过共生体的形成侵入细胞内,共生体在结节细胞的细胞质中释放。然而,hrpG突变体在共生体释放后迅速退化(图3a)。该研究在非共生的原始GMI1000+pRalta菌株中重构了适应性突变hrcV、hrpG和hrpG- efpr,生成了一组具有更高共生能力的近等基因菌株(图a)。研究也表明共生质粒的增益本身就足以激活整个共生反应的19%/10%(上升/下降)(图b)。
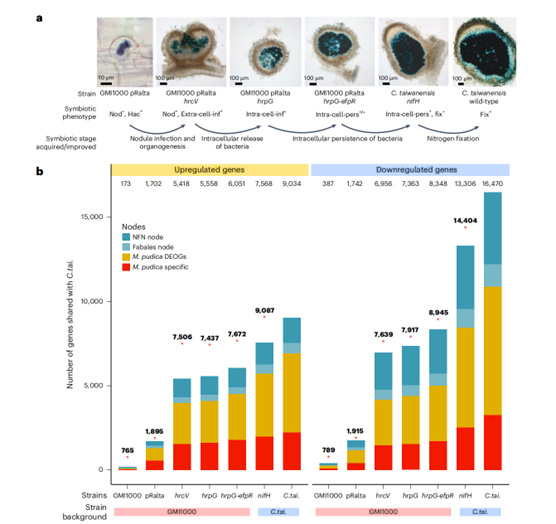
RNS进化实验过程中招募的基因
该研究使用Fisher精确检验来比较整个共生转录组反应和每个性状节点,结果表明,很多对根瘤形成和维持至关重要的转录因子在推断的“器官发生和感染”的祖先转录组特征中被发现。在现有的RNS形成物种中,控制对“Nod因子”、结核“感染和器官发生”和“氮固定”反应的核心机制可能从它们的最新共同祖先开始就被保存下来。
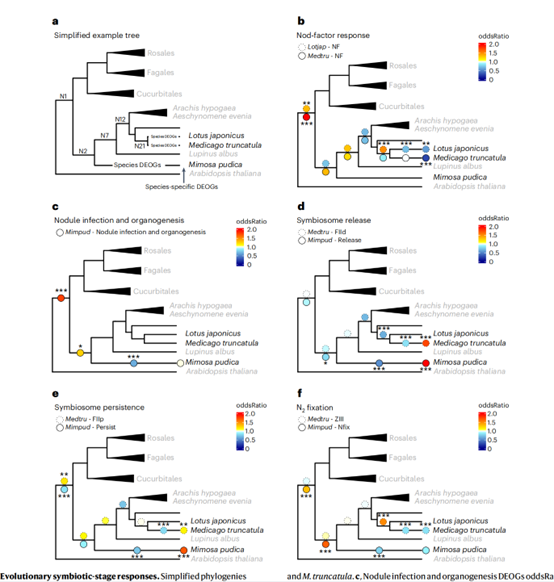
共生进化阶段的反应
最后,该研究以特异性的方式分析了pudica和truncatula中与“共生体释放”和“FIId”相关的上调基因的性质。结果表明进化稳定性是通过共生体释放的趋同进化和细菌共生体控制的增强而获得的,这种控制是通过扩展抗菌肽基因家族获得的。共生体的趋同释放至少由两个独立但类似的分子过程进化而来:已经在dalbergioid和IRLC中提出的结节诱导的小蛋白的进化。
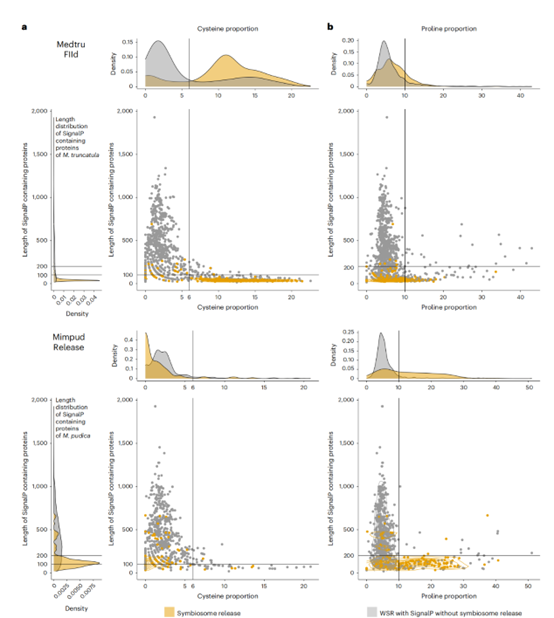
物种特异性DEOGs信号肽蛋白的特征与截断分枝线虫和pudica分枝线虫的共生体释放的相关性
在祖先RNS转录组特征被定义后,未来的研究不得不解释RNS状态是如何从非RNS状态进化而来的。考虑到其与所有细胞内植物共生体的系统发育联系,以及从不同的rns形成的物种中获得的反向遗传数据,可以预测共同共生途径在这一过程中的作用。
原文参考:
https://www.nature.com/articles/s41477-023-01441-w
产品订购:sales@amyjet.com
邮政编码:430070
公司地址:武汉市洪山区光谷大道35号
光谷总部国际二期时代1栋13楼
提示:本公司所有产品仅供科研使用,不用于临床诊断。
版权所有:艾美捷科技有限公司 鄂ICP备10204150号-1 鄂公网安备:42018502004523号
第二类医疗器械经营备案凭证:鄂汉药监械经营备20234324号

微信扫码在线客服