免疫突触(IS)可以定义为免疫细胞在与另一个细胞界面处分子的有序重组(Orange, 2008)。IS存在于抗原呈递细胞(APCs)或靶细胞与淋巴细胞如T细胞、B细胞或自然杀伤细胞之间。这种相互作用对于防御广泛的病原体和异常宿主细胞至关重要。这种相互作用的失调使宿主极易受到病原体的侵害或肿瘤逃逸,另一方面则可能导致自身免疫性疾病(Dustin, 2014)。IS和神经突触共享一些特性,如突触间隙、粘附分子、稳定性和极性,然而免疫细胞之间的相互作用是短暂的,持续时间从几分钟到几小时不等(Friedl et al., 2004)。
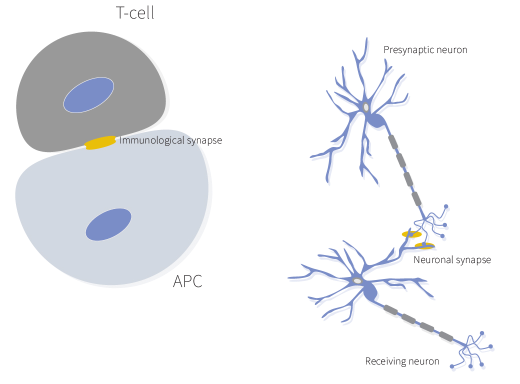
细胞类型特异性的免疫突触
根据所涉及的细胞类型,免疫突触可以分为三种功能亚型(Mastio et al., 2020):溶解性免疫突触(细胞毒性免疫细胞与恶性细胞之间的界面)、抑制性突触(细胞毒性淋巴细胞与健康细胞之间的界面)和调节性突触(细胞毒性细胞与抗原呈递细胞之间的界面,例如树突状细胞)。因此,IS的形成具有许多不同的功能,如配体识别、信号放大和整合、共刺激、细胞毒性、蛋白质分泌和转移、细胞命运决定、激活抑制和信号终止(Orange, 2008年综述),这取决于参与IS形成的细胞类型和受体。
在感染弓形虫的小鼠肝脏中APCs的CD86表达增强
CD86(簇分化86,也称为B7.2)属于B7家族的免疫调节细胞表面蛋白配体,通常在静息B细胞、树突状细胞和巨噬细胞上表达水平较低。激活会导致CD86表达增强(Collins et al., 2005)。CD86和遗传上密切相关的CD80蛋白(也称为B7.1)由抗原呈递细胞表达,通过与T细胞上表达的CD28和细胞毒性T淋巴细胞抗原4(CTLA-4,CD152)相互作用,提供T细胞激活和耐受所需的共刺激信号。CD28的结合导致T细胞激活(Sansom et al. 2000),而CTLA-4的结合则减弱T细胞反应,并作为T细胞激活的负调节器。CD28超家族成员PD-1在与其配体PD-L1结合后也负向调节T细胞激活(Brunner-Weinzierl et al., 2018)。CD80和CD86在免疫调节中扮演非等价的角色:CD86是调节性T细胞(Tregs)增殖和生存的主要配体(Halliday et al., 2020)。与CD80相比,CD86在增强T细胞杀伤能力方面表现出非常高的效率(Thiel et al., 2010)。
弓形虫(Toxoplasma gondii)是一种细胞内原生动物寄生虫,它不仅感染大脑,还感染其他器官,特别是肝脏。肝脏感染的特点是单核炎症细胞的多灶性聚集(Fernández-Escobar et al., 2021)。在小鼠中,弓形虫感染已被证明可以上调CD86的表达,但不影响CD80在单核细胞、巨噬细胞和树突状细胞中的表达(Fischer et al., 1999)。
使用免疫细胞标记物IBA-1、CD86、CD11b和Chil3对未感染对照小鼠和感染弓形虫的小鼠肝脏切片进行免疫组化染色,结果显示在感染弓形虫的小鼠肝脏中IBA1阳性巨噬细胞高度浸润,以及CD86+、CD11b+和Chil3+细胞的多灶性聚集。在未感染对照小鼠中,IBA-1主要染色Kupffer细胞,且很少检测到CD86阳性APCs。未感染肝脏中CD11b和Chil3阳性主要限制在少数CD11b+骨髓源性单核细胞或Chil3+骨髓源性中性粒细胞。
使用小鼠巨噬细胞标记物F4/80和CD11b对肝脏切片进行免疫荧光双重染色,结果显示在未感染和感染弓形虫的小鼠肝脏中均存在F4/80high CD11b-细胞。然而,F4/80low CD11b+细胞仅在感染弓形虫小鼠肝脏的单核细胞聚集中检测到。F4/80+ CD11b-染色识别Kupffer细胞,而F4/80low CD11b+染色特征为在炎症条件下招募的单核源性巨噬细胞(参见:小鼠驻留巨噬细胞)。
免疫荧光双重染色显示,在感染弓形虫的小鼠肝脏单核细胞聚集中,CD86表达主要在F4/80low或F4/80-细胞中增强,这些细胞被特征化为激活的抗原呈递细胞。
通过使用T细胞标记物CD3e(T细胞系标记物)、CD4(T辅助细胞)或CD8a(细胞毒性T细胞)对肝脏切片进行免疫组化染色,可以观察到CD4+ T辅助细胞和,较少程度上,CD8+细胞毒性T细胞大量浸润感染弓形虫的小鼠肝脏。
T细胞免疫已被证明对感染弓形虫的宿主存活至关重要。CD4+ T细胞对早期干扰素-γ的产生很重要。CD8+ T细胞被认为是对抗这种寄生虫的主要效应细胞(Casciotti, 2002)。对感染弓形虫的小鼠肝脏切片进行CD4和CD86的免疫荧光双重染色显示,CD4+ T辅助细胞在单核细胞聚集中与CD86阳性APCs密切接触,形成免疫突触。
T细胞与APCs接触点的超分子组织首次在1998年被可视化(Monks et al., 1998)。免疫突触的关键分子不仅在界面处被覆盖,而且被组织分布在界面的不同区域。这些区域被称为超分子活化复合物(SMACs)(Huppa et al., 2003年综述)。超分子活化复合物的中心区域(cSMAC)在T细胞位点富集T细胞受体(TCRs)和共刺激受体(CD28、CTLA-4),在APC位点富集肽-MHC(pMHC)和B7配体(CD86、CD80)。cSMAC被认为是信号终止和受体循环的位点(Verboogen et al., 2016)。细胞粘附分子如ICAM-1或粘附分子白细胞功能相关分子-1(LFA-1)位于c-SMAC周围的环状结构中,被称为外周SMAC(p-SMAC)。这些细胞粘附分子为免疫突触提供机械支架。大分子如CD43和CD45位于更远端的区域,称为dSMAC。在细胞毒性T细胞和NK细胞中发现的分泌突触中,溶菌颗粒的释放发生在cSMAC的另一个结构域:分泌结构域(Watanabe et al., 2018)。
艾美捷Synaptic Systems:CD86相关产品:
Cat. No. Product Description Application Quantity
HS-384 117 CD11b, rat, monoclonal, purified IgG mouse specific WB IHC IHC-P IHC-Fr 200 ?l
HS-413 108 CD3e, rabbit, monoclonal, recombinant IgG mouse specific IHC IHC-P 100 ?l
HS-360 117 CD4, rat, monoclonal, purified IgG mouse specific WB IHC IHC-P IHC-Fr 200 ?l
HS-466 003 CD86, rabbit, polyclonal, affinity purified mouse specific WB IHC IHC-P 50 ?g
HS-361 003 CD8a, rabbit, polyclonal, affinity purified mouse specific WB IHC IHC-P IHC-Fr 200 ?l
HS-442 017 Chil3 / YM1, rat, monoclonal, purified IgG mouse specific WB ICC IHC IHC-P 100 ?g
HS-397 008 F4/80, rabbit, monoclonal, recombinant IgG WB IHC IHC-P 100 ?l
HS-397 017 F4/80, rat, monoclonal, purified IgG WB IHC IHC-P IHC-Fr 200 ?l
HS-234 017 IBA1, rat, monoclonal, purified IgG WB ICC IHC IHC-P 200 ?l
Synaptic Systems:CD86部分文献:
Orange, 2008. Formation and function of the lytic NK-cell immunological synapse. PMID: 19172692
Collins et al., 2005. The B7 family of immune-regulatory ligands. PMID: 15960813
Dustin 2014. The immunological synapse. PMID: 25367977
Friedl et al., 2004. Diversity in immune-cell interactions: states and funtions of the immunological synapse.
PMID: 15450978
Mastio et al. 2020. Higher Incidence of B Cell Malignancies in Primary Immunodeficiencies: A Combination of Intrinsic Genomic Instability and Exocytosis Defects at the Immunological Synapse. PMID: 33240268
产品订购:sales@amyjet.com
邮政编码:430070
公司地址:武汉市洪山区光谷大道35号
光谷总部国际二期时代1栋13楼
提示:本公司所有产品仅供科研使用,不用于临床诊断。
版权所有:艾美捷科技有限公司 鄂ICP备10204150号-1 鄂公网安备:42018502004523号
第二类医疗器械经营备案凭证:鄂汉药监械经营备20234324号

微信扫码在线客服